泰国斗鱼色氨酸羟化酶基因cDNA在雄鱼好斗性中的关键角色
发表时间: 2023-09-13 21:44

文/纵观人间事
编辑/纵观人间事
泰国斗鱼隶属于硬骨鱼类门鲈形目、攀鲈亚目、丝足鲈科、斗鱼亚科,因其颜色鲜艳、形态多样、易于饲养等特点而成为众多观赏性鱼类中的宠儿。
泰国斗鱼在性别上存在着明显的性别分化,雄鱼具有天然的侵略性和廉价的经济优势,是研究鱼类侵略性的理想试验动物。
进攻性是一个由多个遗传因子共同作用的性状。
研究表明,5-HT通路、多巴胺通路、组胺通路、下丘脑-垂体-肾脏轴、下丘脑-神经-垂体系统、下丘脑-垂体-性腺系统、一氧化氮通路、生长激素statin通路等都参与了该过程。
5-羟色胺又称血清素,由吲哚和乙胺组成,是一种单胺类神经递质,在动物体的情绪调节、体温调节、新陈代谢调节中起重要作用,是5-羟色胺通路中的重要物质之一,其转运体和受体是许多神经类药物的作用靶点。
已有研究表明,5-羟色胺与鱼类攻击性调节有着非常紧密的联系,其参与的信号传导途径中的色氨酸羟化酶(Ttph)在交配竞争后,在交配群体中的分布会发生改变。
而5-羟色胺则会在交配竞争中被显著地上调,从而改变了交配群体的分布格局。
然而,目前对其侵略性的分子调节机理还知之甚少,亟需深入研究。
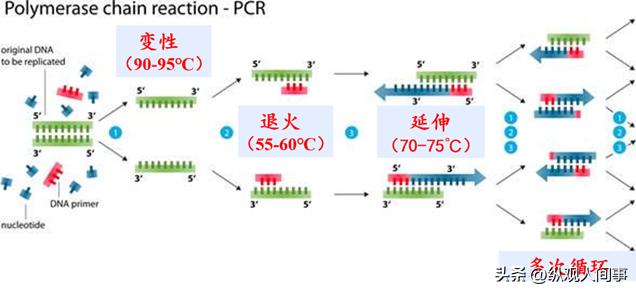
因此,本项目拟采用RT-PCR技术,首先从泰国斗鱼大脑中克隆并鉴定出优秀氨酸羟化酶基因的cDNA片段,其次采用配对对抗和镜像对抗两种方式,观测并纪录泰国公斗鱼的搏杀过程。
随后再采用荧光定量PCR技术,对比对抗过程中各个体之间以及优势与弱势个体之间tphmRNA的表达量变化,从而揭示tph在泰国斗鱼攻击性中的作用机理。
实验用鱼为泰国公斗,体长5-8厘米,可在网店里买到;仪器设备为台式冰冻离心装置、荧光实时定量PCR装置。
通过美国全国生物科技资讯中心,查找与泰国斗鱼相近的鱼的tphcDNA基因。
本项目拟以尼罗罗非鱼、军曹鱼和尖齿胡鲤为研究对象,采用ClustalX软件比较其与其它几种不同类型的tph蛋白家族成员。
通过比较其家族成员之间的差异,设计出相应的简化引物,并结合已有的数据,对泰国斗鱼的tph基因家族成员进行初步的功能验证(表1)。

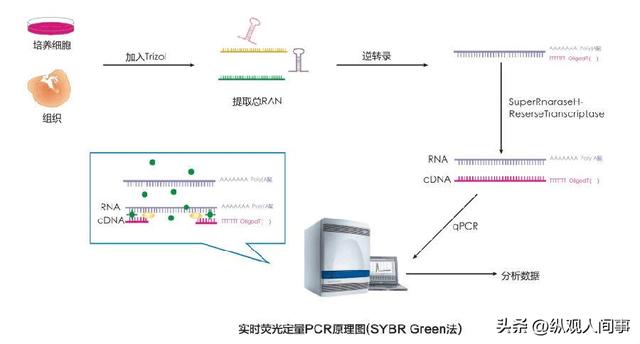
随后应用Trizol技术进行了大鼠的全RNA的提取,用消毒过的剪刀和镊子将泰国斗鱼脑片取出,置于1.5ml的离心机中,加入1ml的LTrizol液体,用一次性针筒搅拌,使其达到均一。
将其置于室温5分钟后,添加0.2毫升三氯甲烷,振荡30秒;在室温条件下,5分钟,4℃,12000r.m-1,离心15分钟;取400ml的上清液于新的1.5ml的离心器中,加0.4ml的异丙醇,将该离心器轻轻翻转几次,然后在室温静置10分钟;4℃,12000r.m-1,离心10分钟后,在其底部发现了一些RNA的白色沉淀物。
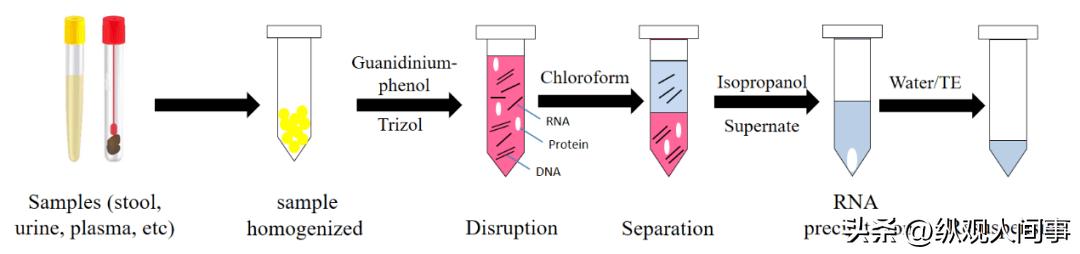
将上清液仔细倒出,再加1mL75%酒精,将沉淀物冲洗干净,重复冲洗两遍;去除上清液,开启瓶盖,使其在室温下自然干燥。
然后再添加10-20微升的脱核糖核酸酶水,使其溶解核糖核酸沉淀;以0.8%的琼脂凝胶电泳法测定RNA的含量,并将其置于-70℃的冷冻环境中进行冷冻保存。
由于mRNA3'端有poly(A)尾,因此以OligdT为引物,通过反转录酶将mRNA反转录到cDNA上。
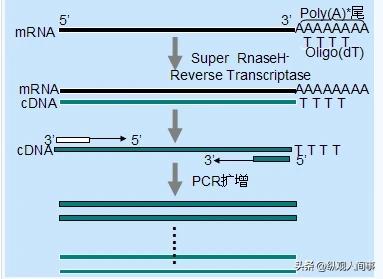
在得到cDNA之后,使用内参考基因beta-actin引物对其进行PCR,然后对PCR产品进行分析。
如果PCR产品条带是单色的,并且是清楚的,那么就说明该产品的品质是合格的,可以作为后续试验的依据。
采用高品质cDNA作为模板,使用表1所示的简并引物对tphcDNA片段进行了扩增。
然后,根据预测的长度,对其进行测序,获得tph基因的某些碱基序列;采用PrimerPrimer5.0开发了特异性的引物,并进行了PCR检测。
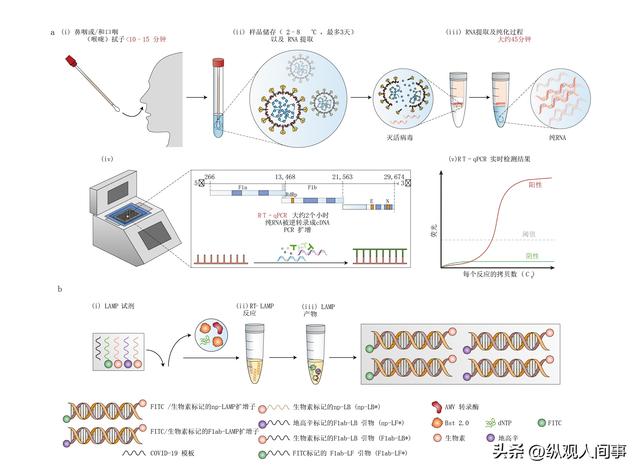
配偶竞争方法是将两种雄性鱼种放在相同的环境中,通过对它们的竞争行为进行观测和记录,根据竞争的最终结果,将它们划分为优势(D)和弱势(I)。
镜象搏斗是指将一块镜子放置在一个大约22.5度的角度下,用它来观测和记录雄性和它的镜象之间的“打斗”。
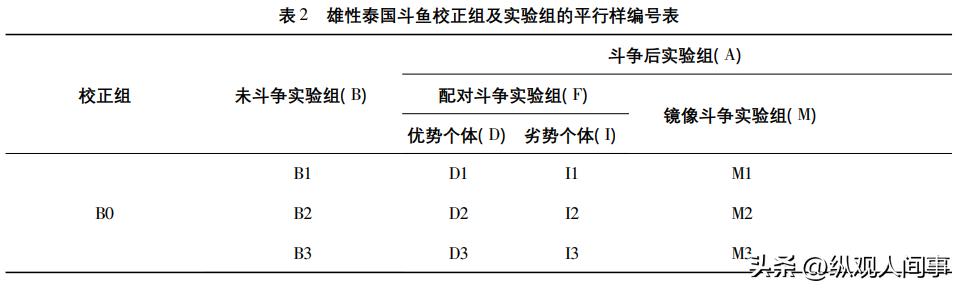
试验的具体方案是选择13条大小与规格相近的泰国公斗鱼,将它们设定为未经斗争的实验组(B)、配对斗争的实验群(F)、镜像斗争的实验群组(M)。
这两条均为后斗争群落(A),每个未经斗争的实验群落(B)需要1条雄的,每个配对斗争群落需要2条公的,每个镜像对抗群落(M)需要一条公的。
每个对照组需要重复3个周期。
为了对定量进行修正,还需要一条未经任何治疗的雄体B0,因此总共需要13条雄体泰国雄体B0(表2)。
纠正后的泰国斗鱼与非抗争模型对照组均不作其他处理,而是取其大脑,抽取其全部RNA。
在进行配对斗争的实验组中,优势个体被记录为D,弱势个体被记录为I,一共展开了3次实验,最终得到了3个实验组D1、I1,D2、I2,D3、I3。
在每一实验组斗争结束之后,都需要快速收集斗鱼的全脑,并提取总RNA。
首先,以修正组B0的cDNA产物为模板,利用特异性引物对进行荧光定量PCR,通过相对定量方法获得不同组分tph的表达水平。
在此基础上,我们使用2-*△Ct方法来处理这些数据,并使用对照样、待检样中的内标基因和目标基因的循环阈值,将内标基因的Ct值作为内源性控制来修正目标基因,从而得到待测样的相对表达量。
将两组间的QR值分别导入SPSS17.0中,通过多因素比较和多因素比较。
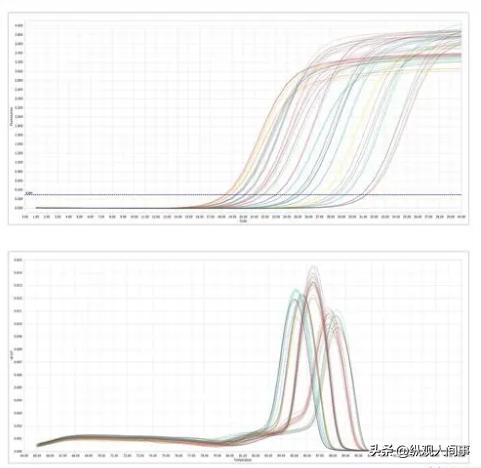
随后使用0.8%琼脂糖凝胶电泳,对各样本总RNA的质量进行检测,将具有良好品质的总RNA作为模板,展开下一步反转录实验,获得cDNA。
以cDNA为模板、β-actin为引物,展开PCR扩增,之后用1.5%琼脂糖凝胶电泳,对PCR产物进行扩增,如果每份cDNA都扩出单一、清晰的β-actin条带,则说明cDNA的质量是良好的(图1)。

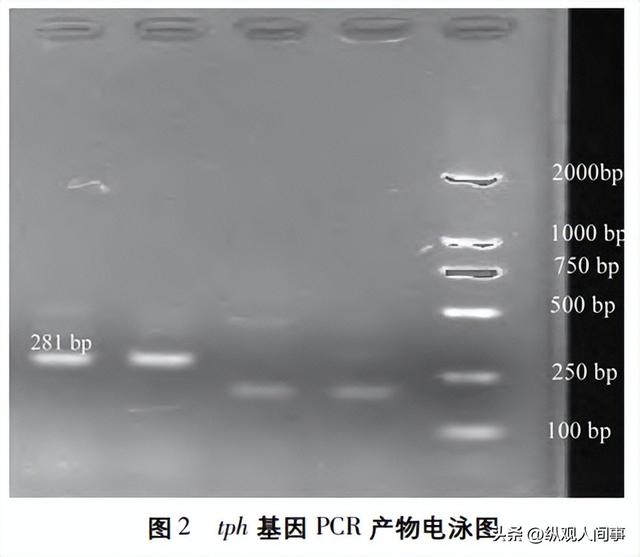
利用tph-F、tph-R两个简化引物,将修正组B0的cDNA作为模板,对tph的cDNA进行扩增,再用1.5%的琼脂凝胶电泳对PCR产物进行测定。
结果显示,281bp附近存在所需的条带(图2)。
利用NCNI的NucleotideBLAST软件对上述基因进行了同源分析,结果表明:该基因与军曹鱼、黄鳝、大黄鱼、黄金鲈、高体鰤、半滑舌鳎、大刺鳅、黄条鰤、尖吻鲈、南极鳕、印度玻璃鱼、牙鲆、深裂眶锯雀鲷中的tph基因具有94%的同源性。
结果表明,所获得的cDNA片段属于泰国斗鱼tph基因的一部分。
将测序所得的核酸序列,输入到DNAStar7.1软件Editseq程序中,人工搜索打开读取框ORF,可以获得84个氨基酸序列,其结果是:
KGELRAYGAGLLSSISELKHALSGNARIMPFDPKVTSKQECIITTFQDVYFVSDSFEEAKVKMREFAKTIKRPFTVRYNPYTQR。
通过对其它一些已知的鱼类tph蛋白的比较,发现泰国斗鱼的tph蛋白与军曹鱼、黄鳝、大黄鱼、黄金鲈、高体鰤、半滑舌鳎、黄条鰤、尖吻鲈等物种的tph蛋白相似度达到97.59%;与刺鳅,牙鲆,深裂眼齿鱼等物种的tph蛋白相似度达到96.39%,显示出较高的物种一致性。
利用NCBI数据库检索其它种类和泌乳纲、爬行纲的几个代表种的tph核酸,并与泰国斗鱼的tph核酸进行比较。
之后再利用MEGA7软件将其转化为可被MEGA7鉴定的形式,并利用邻近比邻法构建进化关系(见图3)。
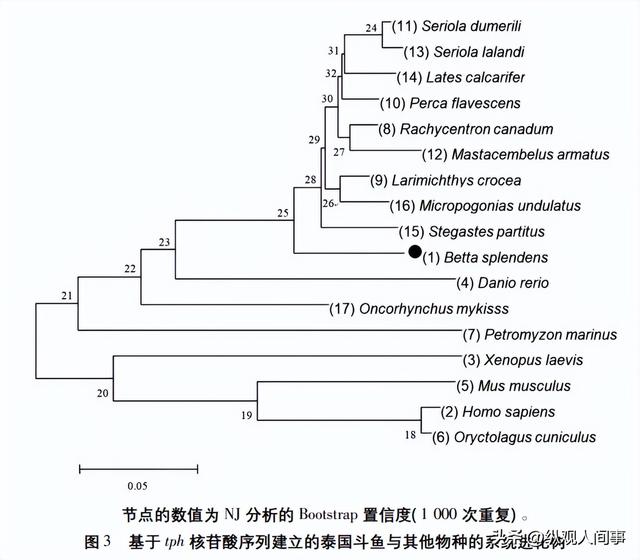
泰国斗鱼与军曹鱼、大黄鱼等均为鲈形目,在演化树中为一个大分支,与鲑形目中的虹鳟鱼、鲤类斑马鱼之间的基因间隔较为遥远,而与海洋七鳃鳗、小白鼠、兔子、智人和非洲爪蛙之间的基因间隔则呈递增趋势。
将两条雄性鱼类同时放在一个透明的烧杯中,双方立刻开始了一场战争。
A会突然做出一个摇晃动作,让迎面而来的B会改变自己的方向,或者是A会突然袭击附近的B,让它逃脱并回到原来的位置。

然后它们就会绕圈,然后就会尝试去咬彼此。
当将镜子放入有一尾雄性斗鱼的透明烧杯中后,斗鱼立刻展现出好斗性行为:腮张开,鳍条直立和身体颤动及起伏等,好斗性强的个体更表现在用尾巴碰撞镜像、张嘴撕咬镜像等斗争行为。
通过对不同个体间tph基因mRNA在抗争前后和抗争后的变化,分析其在抗争前后和抗争后的变化。
试验共分成5组:校对组(B0),斗争之前组(B),配对斗争组(M),其中B组、D组、I组、M组各由3个并行组成。
将测序结果输入到PrimerPrimer5.0软件中,设计出各项得分最高的特异性引物:qtph-F、qtph-R,此引物的溶解曲线呈现为单峰,这证明PCR产物专一,引物选择性强,可以应用于定量表达分析。
将校对组B0作为控制样本,将它的tphmRNA表达值设置为1,对剩余样本的相对表达值QR进行方差分析,结果显示在表3中。

方差分析显示,D组、I组和M组与B组比较有统计学意义(P<0.05);其中,D组和I组与B组比较有显著性意义(P<0.01)(表3)。
上述研究结果显示,不论成对或成群的抗争,其tphmRNA在抗争后的表达量均显著增加;配对斗争中,优势个体D的表达量极显著高于劣势个体I,也极显著高于镜像斗争后个体M。
本文采用RT-PCR方法,从泰国斗鱼体内获得了一个253个碱基片段,对泰国斗鱼tph基因进行了BLAST同源分析,发现其与其它种类的tph基因具有90%的同源性,由此推测该基因可能是泰国斗鱼tph基因的一部分。
通过BLAST的氨基酸序列比较,发现泰国斗鱼tph与军曹鱼、黄鳝、大黄鱼、黄金鲈、高体鰤均有超过96%的相似度,说明其与其它种类的tph基因存在较高的相似性。
通过对泰国斗鱼的分子演化图谱分析,发现其与其他物种之间的基因演化密切相关,而与三文鱼和鲤鱼的基因演化密切相关。
已有报道显示,雄鱼经配对搏斗后,优势种的5-HT系统在末端脑区呈活化状态,而弱势种的5-HT系统则呈活化状态。
通过群体交互作用,我们发现在雌雄斑马鱼中,tph在其优势种群的下丘脑中显著高表达。
与以上侵略性的实验结果类似,不论采用配偶抗争或镜像抗争,泰国斗鱼在抗争后其体内都显著大于非抗争者。
在对战状态下,优势种D的表达量明显大于弱势种I,且在经过对战状态后,也明显大于镜像状态下的M,说明在泰国斗鱼种群处于对战状态时,其tph基因的表达力明显升高。
由此说明tph与雄性泰国斗鱼的好斗性呈明显的正相关,说明色氨酸羟化酶基因tph表达确实参与了泰国斗鱼好斗性的调控,但具体的分子调控机制还有待进一步深入研究。
参考文献:
[1]陈思行,李励年《暹罗斗鱼的饲养与繁殖》
[2]王桂香《骁勇善战的斗鱼》
[3]王婷《拼命将军“泰国斗鱼”》
声明:本站内容部分源于网络转载,出于传递更多信息之目的,并不意味着赞同其观点或证实其描述。文章内容仅供参考,请咨询相关专业人士。
如果无意之中侵犯了您的版权,或有意见、反馈或投诉等情况, 请联系本站,[qq:]
Copyright ©2024 皮球鱼知识网 All rights reserved 版权所有 移动站 鲁ICP备09004228号-92
 鲁公网安备37020202370222号
鲁公网安备37020202370222号