短毛兔与长毛兔皮肤组织学及转录组差异对比研究
发表时间: 2023-08-07 23:55
在阅读此文之前,麻烦您点击一下“关注”,既方便您进行讨论和分享,又能给您带来更多的参与感,感谢您的支持。
文 | 独言
编辑 | 独言

兔子是小型哺乳动物,不仅提供肉,还提供毛发;毛发由毛囊产生,作为哺乳动物特有的皮肤附属物,其特征是周期性再生;兔毛是纺织工业中最受欢迎的天然纤维之一。
影响兔毛经济价值的关键性状包括纤维直径、密度和长度,这由遗传和环境决定。调节头发特征的机制很复杂。了解毛发性状的遗传原理有助于阐明毛发发育的机制,从而促进兔子的繁殖。

毛纤维长度是安哥拉兔毛生产中的一个关键经济特征,因为它与羊毛质量和产量密切相关。此外,头发长度影响纺织品质量,也决定了制造系统。兔子不同类型的毛发具有不同的生长速度,这些生长速度随品种、营养和季节而变化。
在生长期,毛囊通过细胞增殖迅速生长并达到最长的长度;在退行期通过细胞凋亡消退后,它们进入休止期。细胞凋亡是调节毛发周期的关键事件,因为生长期毛发通常在人类中生长4-7年,然后周期进入退行期和休止期的休息阶段。

在毛囊发育过程中,Wnt信号在毛囊形态发生和再生中起着至关重要的作用。此外,一些信号分子,如骨形态发生蛋白、声波刺猬、成纤维细胞生长因子和转化生长因子-β,参与调节毛囊周期进入生长期的时间。
最近的研究表明,VEGFR-22和转录因子NF-κB参与毛发周期调节,NF-κB对于小鼠毛囊干细胞的活化、维持和/或生长是必不可少的。

随着分子生物学技术,特别是二代测序的快速发展,大规模基因表达检测成为可能。兔基因组测序已被用于研究兔驯化过程中表型变异的多基因基础,以及在相同条件下培养的孤雌生殖和正常胚胎中的差异基因表达。
长非编码RNA的景观也已通过RNA测序在毛癣菌诱导的兔皮肤癣菌病病变皮肤和正常皮肤中进行了分析。
在毛发生长的第四周,在短毛和长毛兔中观察到完整的毛囊结构,这表明短毛和长毛兔的毛囊处于生长期。与第1周相比,第1周长毛兔无明显变化;然而,短毛兔的毛囊在第六周出现萎缩。

与第1周相比,初级和次级毛囊面积和内根鞘厚度明显减少。长毛兔的毛囊结构在第八周时保持完整,而短毛兔的毛囊明显萎缩,手指状萎缩至毛发,这表明短毛兔的毛囊在第八周已进入休止期,导致毛发生长停止。然而,长毛兔的毛囊直到第八周仍处于生长期,导致毛发持续生长。
构建了长毛兔组和短毛兔组的36个cDNA文库。去除低质量读数和衔接序列后,每个样品获得超过30 M的干净读数,Q91为≥74.1%。长毛兔和短毛兔测序皮肤RNA的独特读>率为45.2%。

有趣的是,在所有样本中发现的基因数量相对于FPKM值相似。最大比例的基因表现出低和中等表达,一小部分基因表达水平高,进一步证实了通过高通量测序检测低丰度基因的优势。
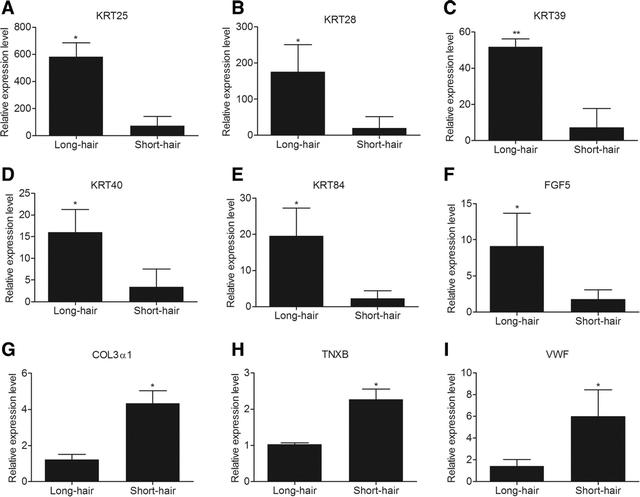
在包括长毛兔皮肤中高表达的0个基因和短毛兔皮肤中高表达的01个基因组成的组之间过滤了2个DEGs。许多被确定为在长毛兔中表达更高的基因来自角蛋白家族,这表明这1个基因是调节兔毛长的重要候选者。
此外,对DEGs进行了主成分分析,结果表明短毛兔和长毛兔在组分2的方向上有明显的区别。此外,PCA鉴定了几种在头发生长调节中起作用的基因。

为了评估我们的DEG文库,通过q-PCR分析了来自KRT家族的五个DEG的相对表达水平以及FGF5和ECM相关基因。总的来说,q-PCR的结果验证了RNA-Seq的结果,并证明了我们数据的可靠性。此外,我们还发现,采摘后第八周所选基因的表达谱与第六周相似。
在本研究中,我们检测到各种剪接事件,包括替代5′第一外显子,替代3′最后外显子,替代外显子末端,内含子保留和跳跃外显子;长毛兔和短毛兔的选择性剪接事件和类型相似。
TSS和TTS发生在5'第一个外显子或3'最后一个外显子的两个或多个剪接位点时,是最常见的观察到的事件,分别约占兔子替代剪接事件的41%和31%。
SKIP是AS中下一个最常见的事件,约占替代剪接事件的17%;IR,即内含子保留在产生的成熟mRNA中,几乎未被检测到,约占AS事件的1.6%。这些结果表明,短毛兔和长毛兔之间每个AS事件的次数相似。
此外,我们使用TopHat2软件将序列数据与已知的参考序列进行比较,然后确定了潜在的SNP。在长毛兔和短毛兔中分别检测到726,852和852,149个SNP。
与长毛兔相比,短毛兔的SNPs更多,表明短毛兔和长毛兔皮肤发育过程中SNPs的差异。大约75%的SNP位点属于转换型,25%的SNP位点属于转型。各样本杂合SNP比例不同,占30.90–34.98%。同时,在每个样本的基因区发现了更多的SNP位点。

短毛兔和长毛兔子在毛发长度方面存在很大的表型差异。毛囊的周期包括三个不同的阶段——生长、消退和休息阶段。这些阶段可以根据毛囊形态进行区分。
在我们之前的研究中,我们通过拔毛重建了兔毛生长的毛囊周期,并分析了短毛和长毛兔的毛发生长节律。
结果表明:长毛兔不同类型毛发从第6周到第31周呈线性上升趋势,短毛兔毛发在9周前呈快速生长,32周后保持稳定,表明两只居群的毛发生长速度和毛发周期不同。

在本研究中,我们在拔毛后第四周、第六周和第八周确定了短毛和长毛兔的毛囊形态。结果表明,毛囊在第八周之前仍处于生长期。
长毛兔毛囊发育的消退期需要进一步研究;短毛兔的毛囊从第四周到第六周开始退化,从第六周到第八周进入休息阶段。毛发生长由膨出的干细胞推动,膨出的干细胞在生长期开始时被真皮激活。

本研究显示,长毛兔毛囊长生长期持续促进毛发生长。在退行期,外根鞘和生发基质中的毛囊干细胞发生凋亡,毛囊下三分之二消退。在休止期,毛囊干细胞相对静止,抑制毛发生长。
这些发现与我们的结果一致,即第六周的毛囊处于退行期,并在拔短毛兔后的第八周进入休止期,导致毛发生长停止。
本研究还提供了一个机会,通过高通量测序比较短毛兔和长毛兔之间的皮肤组织转录组,以分析与毛发生长差异相关的分子机制。获得的转录组数据有助于阐明长毛兔和短毛兔皮肤毛发生长的不同机制。

据报道,每个样品需要>18 M的RNA-Seq读数才能达到饱和状态以进行表达分析。在本研究中,从每个样品中获得了超过36 M的干净读数,并且我们数据的质量控制测定表明RNA-Seq数据是合格的。
对DEGs的热图和PCA的分析显示,在拔毛后第六周,短毛兔和长毛兔的皮肤组织中存在大量显着的DEG。这些基因可能在调节毛囊发育和头发生长中起着至关重要的作用,它们的差异表达可能是短毛兔和长毛兔之间头发长度差异的原因;检测到编码羊毛结构蛋白的10个基因。

KRT的主要功能是保护上皮细胞免受细胞凋亡和可导致细胞死亡的机械或非机械应力。然而,在本研究中,编码角蛋白相关蛋白的几个基因在两个群体中差异表达。
有趣的是,KRT家族10个基因的表达在长毛兔的皮肤组织中上调,这表明KRT基因可能有助于长毛兔毛囊的持续活性。我们假设10个KRT基因也可能有助于促进头发生长。
在早期的报告中,FGF5已被确定为各种哺乳动物毛发生长的抑制剂,FGF5诱导山羊毛囊消退,使用CRISPR/Cas9系统敲除和破坏FGF5导致纤维更长。

然而,有一篇关于兔子的报告表明,FGF5在长羊毛组中也上调,在短羊毛组中也下调,这与我们对RNA-seq和q-PCR的发现一致,表明FGF5 基因在长毛兔的皮肤中上调。
先前的研究还表明,从FGF5 mRNA的替代剪接变体翻译而来的FGF5s多肽抑制了FGF5的活性,增加了头发生长。我们假设FGF5 mRNA的FGF5抑制了FGF5的活性并促进了兔毛的生长。进一步研究需要研究FGF5基因的交替剪接变体在头发长度中的作用。

此外,还选择了这九个基因进行q-PCR验证,以评估我们的DEG文库。所有选定的基因在RNA-Seq和q-PCR之间表现出相似的表达模式,证明了我们数据的可靠性。对所选基因的额外表达分析表明,它们在采摘后第八周的表达谱与第六周相似。
第八周短毛兔和长毛兔的毛囊分别处于休止期和生长期;这一发现进一步表明,这些基因在调节毛囊发育和头发生长中起着重要作用。

此外,还发现许多DEG在细胞外基质受体相互作用和基底细胞癌途径中显着富集。ECM-受体相互作用中的细胞外基质是结构和功能大分子的复杂混合物,包括纤维蛋白。不同的胶原蛋白形成不同的ECM成分,每个细胞的ECM量会影响真皮的体积。
先前的研究表明,许多ECM相关基因在原发性和次级毛囊真皮细胞之间差异表达,导致真皮增大。真皮控制基质细胞的数量,从而控制毛囊及其干的特征和大小。
本研究对COL1α2、COL3α1、COL5α2和COL5α3的差异表达表明,两个群体间毛囊及其干的大小差异与毛囊大小差异有关,PCA进一步表明COL1α2和COL3α1是毛囊发育的重要调节因子。

本研究的结果还表明,参与基底细胞癌通路的GLI1、SHH和PTCH1在短毛兔中的表达上调。据报道,GLI1的抑制诱导细胞周期停滞并增强脑胶质瘤细胞系的凋亡。SHH和PTCH1也是Hedgehog途径的关键组成部分,Hedgehog途径主要调节参与细胞生长、增殖、存活和凋亡的基因。还发现毛囊基因参与湖羊羔羊皮的细胞分化、增殖、凋亡和生长。
因此,GLI1、SHH和PTCH1的差异表达表明了两个群体毛囊周期和细胞凋亡的差异,以及它们在兔毛囊周期和细胞凋亡调节中的作用,WNT信号是毛囊发育启动所必需的。

本研究结果表明,参与WNT信号通路的LEF-1、WNT5a和TCF7在长毛兔中显著上调。此外,Notch信号通路调节哺乳动物耳蜗中的毛细胞分化,并部分依赖于Wnt信号介导毛细胞再生。
在本研究中,我们发现长毛兔TGF-β信号通路中BAMBI和SMAD7的上调;它们是毛发周期的关键调节因子。
我们确定了短毛兔和长毛兔之间皮肤的组织学和转录组谱的差异,组织学分析显示,两个人群的毛发周期和生长速度不同,两个群体转录组谱的差异与形态性状的差异一致。

RNA-Seq使我们能够获得参与毛囊发育,脂质代谢和细胞凋亡的候选基因。兔子皮肤中差异表达的脂质代谢和凋亡基因的表达可用作具有改变毛发长度潜力的新性状。需要进一步的研究来调查候选基因在头发长度中的作用,以改善兔子育种计划。
在麻醉下,在拔毛后的第六周收集背部皮肤样本,此时长毛兔的羊毛周期预计处于生长期,短毛兔的羊毛周期预计处于退行期,立即在液氮中冷冻,并在RNA提取前储存在-80°C。

根据制造商的说明,使用TRIzol试剂从长毛兔和短毛兔的皮肤样品中提取总RNA。使用 Agilent 2100 生物分析仪使用 RNA 完整性数值评估 RNA 的质量。
将mRNA纯化、片段化并转化为cDNA,将接头连接到双链cDNA的末端,并根据制造商的方案使用Illumina TruSeq RNA样品制备试剂盒通过聚合酶链反应创建文库。
声明:本站内容部分源于网络转载,出于传递更多信息之目的,并不意味着赞同其观点或证实其描述。文章内容仅供参考,请咨询相关专业人士。
如果无意之中侵犯了您的版权,或有意见、反馈或投诉等情况, 请联系本站,[qq:]
Copyright ©2024 皮球鱼知识网 All rights reserved 版权所有 移动站 鲁ICP备09004228号-92
 鲁公网安备37020202370222号
鲁公网安备37020202370222号